Pahoitteluni blogin hitaasti päivitystahdista, mutta varsinainen tutkimustyö sekä kesäloma ovat antaneet muuta puuhasteltavaa. Nyt kuitenkin voimme palata blogin pariin virkein mielin.
Edellisissä kirjoituksissa selvitin populaatiogenetiikan perusteita sekä kelpoisuuden käsitettä. Ennen kuin siirrymme luonnonvalinnan tarkasteluun on syytä selventää hieman ekologian perusteita. Tämä toivottavasti valoittaa kelpoisuuden käsitettä hieman lisää.
Oletetaan, että tarkasteltava populaatio lisääntyy suvuttomasti jakautumalla, eli kyseeseen tulee jonkinlainen mikrobi. Olkoon populaation koko  ajanhetkellä
ajanhetkellä  . Ajatellaan, että voimme tarkkailla yhtä populaation yksilöä lyhyen aikaa
. Ajatellaan, että voimme tarkkailla yhtä populaation yksilöä lyhyen aikaa  . Oikeasti mikrobit ovat liian pieniä, jotta voisimme näin tehdä, mutta blogin lukijoiden mielikuvitus varmaankin riittää tähän tehtävään. Olkoon todennäköisyys sille, että tarkkailtava yksilö jakautuu kun tarkkailemme sitä
. Oikeasti mikrobit ovat liian pieniä, jotta voisimme näin tehdä, mutta blogin lukijoiden mielikuvitus varmaankin riittää tähän tehtävään. Olkoon todennäköisyys sille, että tarkkailtava yksilö jakautuu kun tarkkailemme sitä 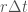 . Oletamme tässä että
. Oletamme tässä että  on vakio, näin ei varmaankaan todellisuudessa ole, mutta yksinkertaistamme hieman keskustelua. Nyt muutos populaatiokoossa on
on vakio, näin ei varmaankaan todellisuudessa ole, mutta yksinkertaistamme hieman keskustelua. Nyt muutos populaatiokoossa on 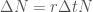 . Kun
. Kun 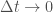 , voidaan muutos populaatiokoossa kirjoittaa muotoon
, voidaan muutos populaatiokoossa kirjoittaa muotoon
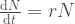
Tämän yhtälön ratkaisu on  , jossa
, jossa  on populaatiokoko ajanhetkellä
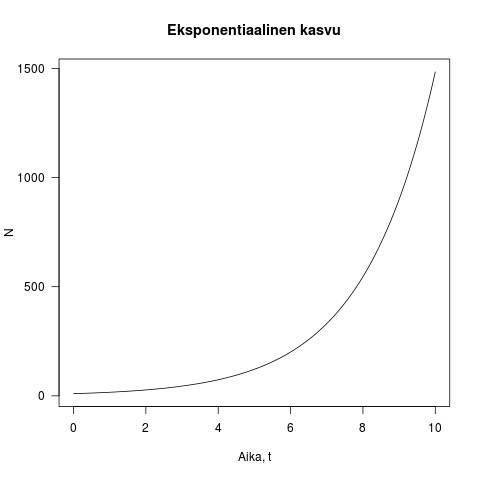
on populaatiokoko ajanhetkellä  . Tämä on eksponentiaalisen kasvun yhtälö. Kun populaatio on pieni, kasvu on ensin hidasta, mutta kiihtyy mitä suuremmaksi populaatio kasvaa (katso kuva 1).
. Tämä on eksponentiaalisen kasvun yhtälö. Kun populaatio on pieni, kasvu on ensin hidasta, mutta kiihtyy mitä suuremmaksi populaatio kasvaa (katso kuva 1).
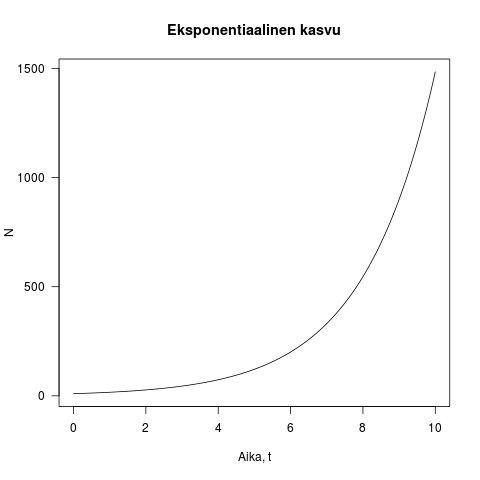
Kuva 1. Esimerkki eksponentiaalisesta kasvusta. Populaation koko on y-akselilla ja aika x-akselilla. Kuvassa  ja
ja  .
.
Oikeasti mikään populaatio ei voi kasvaa eksponentiaalisesti kovinkaan kauaa, sillä ravinteet tai tila loppuvat nopeasti kesken. Tämän voi huomata siitä, että emme ole hukkuneet bakteerimassaan vaikka labrassa ravinnerikkaassa liuoksessa kasvatetut bakteerit voivat jakaantua hyvinkin nopeasti. Kun ravinteet loppuvat kasvun täytyy loppua. Luonnolliset populaatiot voivat pysyä vain tasolla johon ympäristön resurssit riittävät. Tämän huomion teki aikoinaan ensimmäisenä T. R. Malthus 1798 kirjassaan ”An essay on the principle of population”.
Avatkaamme tätä ajatusta hieman lisää. Muutos populaation koossa on syntyneitten yksilöiden ja kuolleiden yksilöiden erotus, 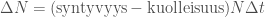 . Oletetaan, että kuolleisuus on vakio, mutta syntyvyys riippuu populaation koosta. Olkoon syntyvyys
. Oletetaan, että kuolleisuus on vakio, mutta syntyvyys riippuu populaation koosta. Olkoon syntyvyys 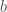 , kuolleisuus
, kuolleisuus 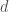 ja kerroin joka kertoo tiheyden vaikutuksen syntyvyyteen
ja kerroin joka kertoo tiheyden vaikutuksen syntyvyyteen 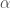 . Nyt voimme kirjoittaa populaation kasvua kuvaavan yhtälön muotoon
. Nyt voimme kirjoittaa populaation kasvua kuvaavan yhtälön muotoon
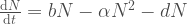
populaation kasvua kuvaava termi on nyt  . Järjestetään termit uudelleen niin saamme
. Järjestetään termit uudelleen niin saamme
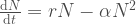
Tässä esimerkissä kuvaa vain syntyvyyden hidastumista, kun tila tai resurssit käyvät vähiin. Voisimme kuitenkin sisällyttää siihen myös populaatiotiheyden vaikutukset kuolleisuuteen. Ratkaisu edellä kuvatulle yhtälölle on

Tätä kutsutaan logistisen kasvun yhtälöksi. Lukijat ovat saattaneet törmätä siihen sellaisessa muodossa, jossa  , termiä
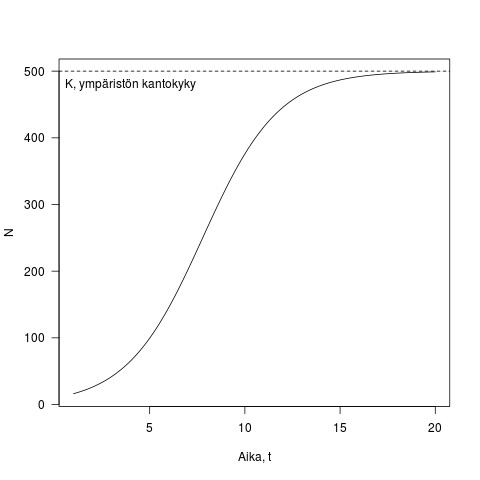
, termiä  nimitetään usein ympäristön kantakyvyksi. Olen esittänyt tämän yhtälön muodossa jossa K:lle on annettu mekanistinen perusta, näimme kohta tämän esitystavan hyödyt. Kuvassa 2 on esimerkki logistisesta kasvusta, populaation kasvu on ensin nopeaa, mutta tiheyden kasvaessa kasvu hidastuu kunnes lopulta pysäähtyy kun populaatio saavuttaa ympäristön kantokyvyn.
nimitetään usein ympäristön kantakyvyksi. Olen esittänyt tämän yhtälön muodossa jossa K:lle on annettu mekanistinen perusta, näimme kohta tämän esitystavan hyödyt. Kuvassa 2 on esimerkki logistisesta kasvusta, populaation kasvu on ensin nopeaa, mutta tiheyden kasvaessa kasvu hidastuu kunnes lopulta pysäähtyy kun populaatio saavuttaa ympäristön kantokyvyn.
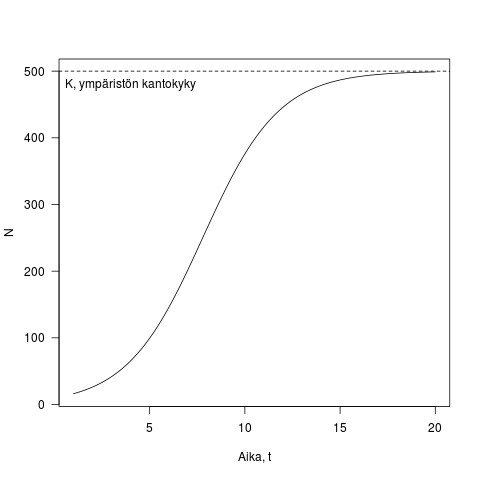
Kuva 2. Esimerkki logistisesta kasvusta. Populaation koko on y-akselilla ja aika x-akselilla. Kuvassa , ja  . Joten
. Joten 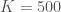 .
.
Luettuaan Malthusia, Darwin ja Wallace tajusivat, että rajallisista resursseista täytyy syntyä kilpailua. Yksilöt jotka pärjäävät kilpailussa parhaiten jättävät (keskimäärin) enemmän jälkeläisiä. Jos kilpailussa pärjääminen on perinnöllistä, tämän tyyppiset yksilöt yleistyvät populaatiossa. Tästä luonnonvalinnassa on kysymys.
Ajatellaan, että kaksi erilaista suvuttomasti lisääntyvää yksilöä, x ja y, kilpailevat samoista resursseista. Näin ollen molempien tyyppien tiheydet vaikuttavat niiden syntyvyyteen. X ja Y ovat erilaisten tyyppien populaatiokoot. Näiden kasvua kuvaavat yhtälöt ovat

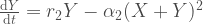
Niinpä  kun
kun  ja vastaavasti tyypille y,
ja vastaavasti tyypille y, 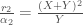 . Kun eri tyypit kilpailevat keskenään se tyyppi jolla on suurempi
. Kun eri tyypit kilpailevat keskenään se tyyppi jolla on suurempi  syrjäyttää toisen. Sillä jos
syrjäyttää toisen. Sillä jos 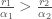 niin populaatio kasvaa kunnes se saavuttaa pisteen jossa
niin populaatio kasvaa kunnes se saavuttaa pisteen jossa 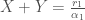 , mutta nyt
, mutta nyt  joten
joten  . Niinpä tyypin y populaatio pienenee ja häviää lopulta kokonaan.
. Niinpä tyypin y populaatio pienenee ja häviää lopulta kokonaan.
Näitä yhtälöitä tarkastelemalla voimme ymmärtää miksi muutokset sekä syntyvyydessä, että kuolleisuudessa voivat muuttaa yksilöiden kelpoisuutta. Oletimme edellä, että termiin sisältyi vain populaatiotiheyden vaikutukset syntyvyyteen. Biologinen tulkinta tälle voisi olla esimerkiksi miten yksilöt kilpailevat resursseista tai tilasta. Voimme kuitenkin sisällyttää tähän termiin myös tiheyden vaikutukset kuolleisuuteen. Biologisesti tämä voisi tarkoittaa esimerkiksi sitä, että kuolleisuus kasvaa populaation tiheyden kasvaessa, koska taudit leviävät helpommin. Termiin sisältyi sekä syntyvyys, että kuolleisuus. Niinpä monet eri tekijät voivat muuttaa yksilöiden kelpoisuutta, kuten jo aiemmassa blogissa todettiin. Toivottavasti tämä esitys valotti asiaa hieman lisää.
Nyt hallitsemme ekologian ja populaatiogenetiikan perusteet siten, että seuraavaksi voimmekin ryhtyä tarkastelemaan itse luonnonvalintaa!
Näihin kuviin näihin tunnelmiin!
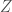 . Ensimmäisessä tapauksessa geneettistä muuntelua on vain yhdessä lokuksessa, jossa on kaksi eri alleelia
. Ensimmäisessä tapauksessa geneettistä muuntelua on vain yhdessä lokuksessa, jossa on kaksi eri alleelia 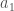 ja
ja  joiden vaikutukset fenotyyppiin
joiden vaikutukset fenotyyppiin  ja
ja 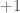 . Oletetaan vielä yksinkertaisuuden vuoksi, että molempien alleelien frekvenssit ovat
. Oletetaan vielä yksinkertaisuuden vuoksi, että molempien alleelien frekvenssit ovat  ja että alleelit ovat Hardy-Weinberg tasapainossa. Tarkastellaan diploideja otuksia, niinpä populaatiossa esiintyy kolmea eri fenotyyppiä:
ja että alleelit ovat Hardy-Weinberg tasapainossa. Tarkastellaan diploideja otuksia, niinpä populaatiossa esiintyy kolmea eri fenotyyppiä: 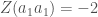 ,
,  ja
ja 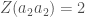 . Tämä tilanne on esitetty kuvassa 1A.
. Tämä tilanne on esitetty kuvassa 1A.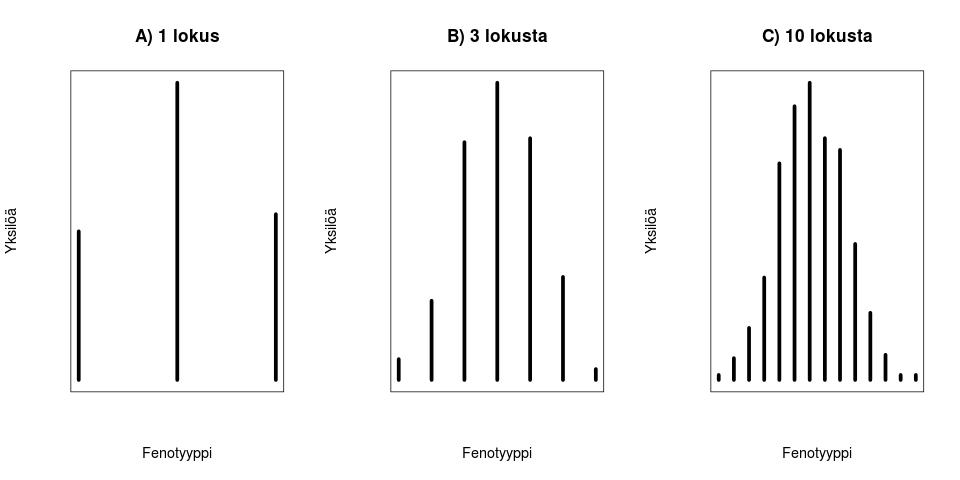
 fenotyyppiä
fenotyyppiä  , jossa
, jossa  on lokuksen
on lokuksen  genotyypin vaikutus fenotyyppiin
genotyypin vaikutus fenotyyppiin 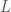 on lokusten lukumäärä ja
on lokusten lukumäärä ja  . Eli toisin sanoen yksilön fenotyyppi määräytyy summaamalla kaikkien fenotyyppiin vaikuttavien lokusten genotyyppien vaikutukset ja lisäämällä satunnaisesta ympäristönvaihtelusta johtuva virhetermi. Tässä oletamme, että satunnaisvaihtelu on normaalisti jakautunut keskihajonnalla
. Eli toisin sanoen yksilön fenotyyppi määräytyy summaamalla kaikkien fenotyyppiin vaikuttavien lokusten genotyyppien vaikutukset ja lisäämällä satunnaisesta ympäristönvaihtelusta johtuva virhetermi. Tässä oletamme, että satunnaisvaihtelu on normaalisti jakautunut keskihajonnalla 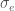 .
.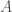 , jolla on alleelit
, jolla on alleelit 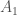 ja
ja  . Hardy-Weinberg tasapaino pätee tuotettuihin jälkeläisiin, mutta ei aikuisiin, koska eri alleeleja kantavat yksilöt selviytyvät aikuisuuteen eri todennäköisyyksillä. Merkitään alleelin
. Hardy-Weinberg tasapaino pätee tuotettuihin jälkeläisiin, mutta ei aikuisiin, koska eri alleeleja kantavat yksilöt selviytyvät aikuisuuteen eri todennäköisyyksillä. Merkitään alleelin 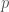 :llä, joten alleelin
:llä, joten alleelin 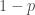 . Voimme taulukoida genotyyppien kelpoisuudet ja frekvenssit seuraavasti.
. Voimme taulukoida genotyyppien kelpoisuudet ja frekvenssit seuraavasti.
 . Miten tämä muuttuu kun
. Miten tämä muuttuu kun 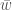 :n yhtälö
:n yhtälö 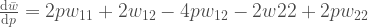
 alleelin
alleelin  ja
ja  .
. . Voimme myös järjestää muutoksen populaation keskimääräisessä kelpoisuudessa uudelleen
. Voimme myös järjestää muutoksen populaation keskimääräisessä kelpoisuudessa uudelleen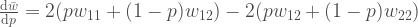

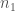

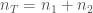 kaikkien alleelien lukumäärä populaatiossa. Näin ollen
kaikkien alleelien lukumäärä populaatiossa. Näin ollen  . Seuraavassa sukupolvessa alleelin
. Seuraavassa sukupolvessa alleelin  ja kaikkien alleelien lukumäärä
ja kaikkien alleelien lukumäärä  . Niinpä p on seuraavassa sukupolvessa:
. Niinpä p on seuraavassa sukupolvessa:

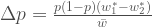
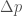 niin saamme:
niin saamme:
 on aina positiivinen tai nolla, joten evoluution suunta (kasvaako vai pieneneekö
on aina positiivinen tai nolla, joten evoluution suunta (kasvaako vai pieneneekö  funktiona
funktiona 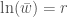 eli populaation kasvua kuvaava kerroin, joten populaatiot aina kiipeävät korkeampaa kelpoisuutta kohti. Kaikki tasapainotilat ovat
eli populaation kasvua kuvaava kerroin, joten populaatiot aina kiipeävät korkeampaa kelpoisuutta kohti. Kaikki tasapainotilat ovat