Pahoitteluni blogin hitaasti päivitystahdista, mutta varsinainen tutkimustyö sekä kesäloma ovat antaneet muuta puuhasteltavaa. Nyt kuitenkin voimme palata blogin pariin virkein mielin.
Edellisissä kirjoituksissa selvitin populaatiogenetiikan perusteita sekä kelpoisuuden käsitettä. Ennen kuin siirrymme luonnonvalinnan tarkasteluun on syytä selventää hieman ekologian perusteita. Tämä toivottavasti valoittaa kelpoisuuden käsitettä hieman lisää.
Oletetaan, että tarkasteltava populaatio lisääntyy suvuttomasti jakautumalla, eli kyseeseen tulee jonkinlainen mikrobi. Olkoon populaation koko 


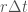

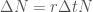
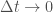
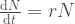
Tämän yhtälön ratkaisu on 


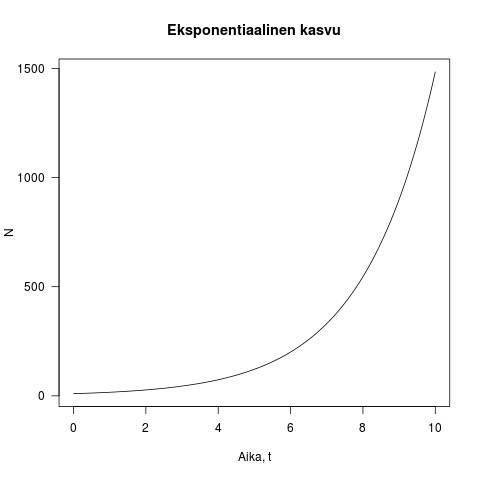
Kuva 1. Esimerkki eksponentiaalisesta kasvusta. Populaation koko on y-akselilla ja aika x-akselilla. Kuvassa 

Oikeasti mikään populaatio ei voi kasvaa eksponentiaalisesti kovinkaan kauaa, sillä ravinteet tai tila loppuvat nopeasti kesken. Tämän voi huomata siitä, että emme ole hukkuneet bakteerimassaan vaikka labrassa ravinnerikkaassa liuoksessa kasvatetut bakteerit voivat jakaantua hyvinkin nopeasti. Kun ravinteet loppuvat kasvun täytyy loppua. Luonnolliset populaatiot voivat pysyä vain tasolla johon ympäristön resurssit riittävät. Tämän huomion teki aikoinaan ensimmäisenä T. R. Malthus 1798 kirjassaan ”An essay on the principle of population”.
Avatkaamme tätä ajatusta hieman lisää. Muutos populaation koossa on syntyneitten yksilöiden ja kuolleiden yksilöiden erotus, 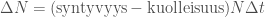
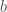
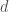
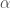
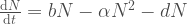
populaation kasvua kuvaava termi 
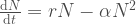
Tässä esimerkissä

Tätä kutsutaan logistisen kasvun yhtälöksi. Lukijat ovat saattaneet törmätä siihen sellaisessa muodossa, jossa 

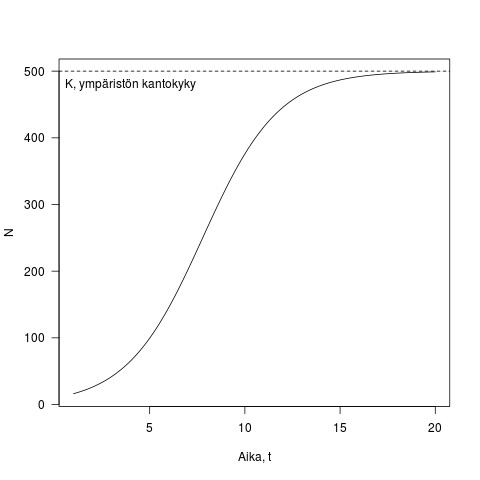
Kuva 2. Esimerkki logistisesta kasvusta. Populaation koko on y-akselilla ja aika x-akselilla. Kuvassa 
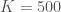
Luettuaan Malthusia, Darwin ja Wallace tajusivat, että rajallisista resursseista täytyy syntyä kilpailua. Yksilöt jotka pärjäävät kilpailussa parhaiten jättävät (keskimäärin) enemmän jälkeläisiä. Jos kilpailussa pärjääminen on perinnöllistä, tämän tyyppiset yksilöt yleistyvät populaatiossa. Tästä luonnonvalinnassa on kysymys.
Ajatellaan, että kaksi erilaista suvuttomasti lisääntyvää yksilöä, x ja y, kilpailevat samoista resursseista. Näin ollen molempien tyyppien tiheydet vaikuttavat niiden syntyvyyteen. X ja Y ovat erilaisten tyyppien populaatiokoot. Näiden kasvua kuvaavat yhtälöt ovat

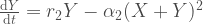
Niinpä 

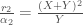

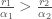
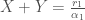


Näitä yhtälöitä tarkastelemalla voimme ymmärtää miksi muutokset sekä syntyvyydessä, että kuolleisuudessa voivat muuttaa yksilöiden kelpoisuutta. Oletimme edellä, että termiin
Nyt hallitsemme ekologian ja populaatiogenetiikan perusteet siten, että seuraavaksi voimmekin ryhtyä tarkastelemaan itse luonnonvalintaa!
Näihin kuviin näihin tunnelmiin!
