Tervehdys jälleen rakkaat lukijat!
Monet teistä ovat odottaneet uutta blogipäivitystä malttamattomina. Aurinkokaan ei ole paistanut niin kirkkaasti ja leipäkään ei ole maistunut leivältä blogipäivityksien puutteessa. Pahoittelen, että en ehtinyt päivittää viimeviikolla, sillä oli kiireitä itse tutkimuksen parissa.
Evoluution määritelmä
Olette ehtineet varmasti sulatella edellistä kirjoitusta. Opimme viimeksi, mitä alleelit ovat. Seuraavaksi aloitamme varsinaisen evoluutioteorian tarkastelun. Monet lukijat varmaan muistavat koulusta erään evoluution määritelmän: ”Evoluutio on alleelifrekvenssien muutosta populaatiossa.” Alleelifrekvensseillä tarkoitetaan eri alleelien osuuksia populaatiossa. Oletetaan, että populaation koko, 
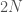

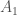

Evoluutiossa on siis kyse siitä, että 
![[0, 1]](https://s0.wp.com/latex.php?latex=%5B0%2C+1%5D&bg=ffffff&fg=444444&s=0&c=20201002)
Genotyyppifrekvenssi
Ennen varsinaista keskustelua kelpoisuudesta, on syytä sanoa muutama sana genotyyppien frekvensseistä. Oletamme tässä, että populaation yksilöt pariutuvat satunnaisesti tarkasteltavien alleelien suhteen. Olkoon populaatiossa on kaksi alleelia, 



Kelpoisuuden käsitteestä
Jälleen lukijat varmasti muistavat koulusta jonkinlaisen luonnonvalinnan toiminnan kuvauksen. Jotkut yksilöt saavat enemmän jälkeläisiä kuin toiset ja tämän seurauksena seuraavassa sukupolvessa on suhteessa enemmän näiden yksilöiden jälkeläisiä. Haluamme kuitenkin määritellä luonnonvalinnan tarkemmin. Ensin määrittelemme kelpoisuuden (engl. fitness), evoluutiobiologiassa on joitakin käsitteitä joilla on tutkimuskäytössä tarkat määritelmät, mutta joita valitettavasti pahoinpidellään julmasti kansan keskuudessa. Kelpoisuus on yksi näistä. Kelpoisuus, 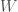
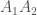
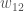
Voimme mitata genotyyppistäkelpoisuutta joko absoluuttisesti tai suhteellisesti. Absoluuttinenkelpoisuus on odotettu eloonjäävien jälkeläisten määrä jotka tietyn genotyypin yksilö tuottaa. Suhteellinenkelpoisuus on absoluuttinen kelpoisuus skaalattuna jollakin tavalla. Usein esimerkiksi siten, että suurin kelpoisuus saa arvon 1. Suhteellinen kelpoisuus on usein hyödyllinen evoluution mallintamisessa, mutta absoluuttista kelpoisuutta käyttämällä näemme miten kelpoisuuden käsite on kytköksissä ekologiaan.
Tarkastellaan populaation kasvua yhden sukupolven verran. Olkoon populaatiokoko ajassa t, 
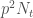
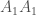



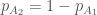
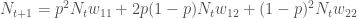
Tämän voimme kirjoittaa muotoon 

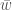
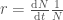
Jos populaation kasvun on hidasta, niin diskreetti- ja jatkuvamalli ovat likimäärin samat, ja näin löydämme yhteyden populaation kelpoisuuden ja ekologiassa käytetyn populaation kasvun kesken. Siis 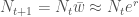
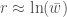
Näin huomaamme, että kelpoisuus siis kytkeytyy ekologiseen teoriaan ja populaation kasvuun. Niinkuin nykyään on tapana sanoa: Fantastista! Kelpoisuudella ei siis ole mitään tekemistä ”vain vahvimmat selviytyvät” tyyppisten sanontojen kanssa. Kyse on siitä miten eri yksilöt vaikuttavat populaation kasvuun. Tietysti yksi osa kelpoisuutta voi olla eloonjääminen, mutta niin ei tarvitse olla.
Kirjallisuutta
Rice, S. H. 2004. Evolutionary Theory: Mathematical and Conceptual Foundations. Sinauer Associates Inc., Sunderland, MA.
