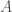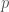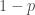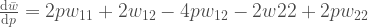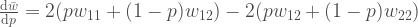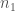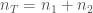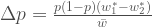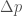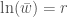Tervehdys jälleen rakkaat lukijat!
Monet teistä ovat odottaneet uutta blogipäivitystä malttamattomina. Aurinkokaan ei ole paistanut niin kirkkaasti ja leipäkään ei ole maistunut leivältä blogipäivityksien puutteessa. Pahoittelen, että en ehtinyt päivittää viimeviikolla, sillä oli kiireitä itse tutkimuksen parissa.
Evoluution määritelmä
Olette ehtineet varmasti sulatella edellistä kirjoitusta. Opimme viimeksi, mitä alleelit ovat. Seuraavaksi aloitamme varsinaisen evoluutioteorian tarkastelun. Monet lukijat varmaan muistavat koulusta erään evoluution määritelmän: ”Evoluutio on alleelifrekvenssien muutosta populaatiossa.” Alleelifrekvensseillä tarkoitetaan eri alleelien osuuksia populaatiossa. Oletetaan, että populaation koko,  , on 100 yksilöä. Oletetaan vielä, että nämä yksilöt ovat diploideja (kuten esim. ihmiset ja muut eläimet). Tästä seuraa, että kyseisessä populaatiossa on
, on 100 yksilöä. Oletetaan vielä, että nämä yksilöt ovat diploideja (kuten esim. ihmiset ja muut eläimet). Tästä seuraa, että kyseisessä populaatiossa on 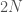 kopiota jokaisesta lokuksesta. Jos populaatiossa on
kopiota jokaisesta lokuksesta. Jos populaatiossa on  alleelin
alleelin 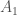 kopiota, alleelin frekvenssi on silloin
kopiota, alleelin frekvenssi on silloin

Evoluutiossa on siis kyse siitä, että  muuttuu sukupolvien myötä. Ei oikeastaan mistään sen mystisemmästä. Huomaa, että voi saada arvoja vain väliltä
muuttuu sukupolvien myötä. Ei oikeastaan mistään sen mystisemmästä. Huomaa, että voi saada arvoja vain väliltä ![[0, 1]](https://s0.wp.com/latex.php?latex=%5B0%2C+1%5D&bg=ffffff&fg=444444&s=0&c=20201002) . Evoluutio ei kuitenkaan pysähdy jos jokin alleeli yleistyy populaatiossa frekvenssiin 1. Mutaatiot tuottavat uusi alleeleja ja vanhat alleelit korvautuvat uusilla evoluution kuluessa. Luonnonvalinta on tärkein alleelifrekvenssejä muuttava voima. Seuraavaksi selvennämme joitakin käsitteitä, jotka ovat välttämättömiä luonnonvalinnan ymmärtämiseksi.
. Evoluutio ei kuitenkaan pysähdy jos jokin alleeli yleistyy populaatiossa frekvenssiin 1. Mutaatiot tuottavat uusi alleeleja ja vanhat alleelit korvautuvat uusilla evoluution kuluessa. Luonnonvalinta on tärkein alleelifrekvenssejä muuttava voima. Seuraavaksi selvennämme joitakin käsitteitä, jotka ovat välttämättömiä luonnonvalinnan ymmärtämiseksi.
Genotyyppifrekvenssi
Ennen varsinaista keskustelua kelpoisuudesta, on syytä sanoa muutama sana genotyyppien frekvensseistä. Oletamme tässä, että populaation yksilöt pariutuvat satunnaisesti tarkasteltavien alleelien suhteen. Olkoon populaatiossa on kaksi alleelia, ja  , joiden frekvenssit ovat ja
, joiden frekvenssit ovat ja  . Nyt genotyyppien frekvenssit ovat
. Nyt genotyyppien frekvenssit ovat  ja
ja  . Oikeastaan olisin ennen tätä kirjoitusta voinut selittää tarkemmin miksi näin on. Nyt pyydän lukijoilta malttia ja vain ottamaan tämän annettuna. Tulen selittämään tämän tulevassa kirjoituksessa. Nokkelimmat lukijat ehkäpä muistavat koulusta, että tähän liittyy jotenkin Mendel sekä herrat Hardy ja Weinberg.
. Oikeastaan olisin ennen tätä kirjoitusta voinut selittää tarkemmin miksi näin on. Nyt pyydän lukijoilta malttia ja vain ottamaan tämän annettuna. Tulen selittämään tämän tulevassa kirjoituksessa. Nokkelimmat lukijat ehkäpä muistavat koulusta, että tähän liittyy jotenkin Mendel sekä herrat Hardy ja Weinberg.
Kelpoisuuden käsitteestä
Jälleen lukijat varmasti muistavat koulusta jonkinlaisen luonnonvalinnan toiminnan kuvauksen. Jotkut yksilöt saavat enemmän jälkeläisiä kuin toiset ja tämän seurauksena seuraavassa sukupolvessa on suhteessa enemmän näiden yksilöiden jälkeläisiä. Haluamme kuitenkin määritellä luonnonvalinnan tarkemmin. Ensin määrittelemme kelpoisuuden (engl. fitness), evoluutiobiologiassa on joitakin käsitteitä joilla on tutkimuskäytössä tarkat määritelmät, mutta joita valitettavasti pahoinpidellään julmasti kansan keskuudessa. Kelpoisuus on yksi näistä. Kelpoisuus, 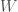 , on yksilön jälkeläisten lukumäärä seuraavassa sukupolvessa. Populaatiogenetiikassa käytämme usein genotyypin kelpoisuutta, joka on kaikkien populaation saman genotyypin yksilöiden keskimääräinen kelpoisuus. Eli genotyypin
, on yksilön jälkeläisten lukumäärä seuraavassa sukupolvessa. Populaatiogenetiikassa käytämme usein genotyypin kelpoisuutta, joka on kaikkien populaation saman genotyypin yksilöiden keskimääräinen kelpoisuus. Eli genotyypin 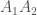 genotyyppinenkelpoisuus,
genotyyppinenkelpoisuus, 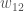 , on kaikkien niiden yksilöiden :n keskiarvo, joilla on genotyyppi . Usein populaatiogenetiikassa oletamme, että luonnonvalinta kohdistuu yksilötasolla fenotyyppeihin ja nämä fenotyypit määrää yksilön genotyyppi. Niinpä kelpoisuuden määritelmä yksilön ominaisuudeksi ei ole ongelma vaikka sovellamme kelpoisuutta genotyyppeihin.
, on kaikkien niiden yksilöiden :n keskiarvo, joilla on genotyyppi . Usein populaatiogenetiikassa oletamme, että luonnonvalinta kohdistuu yksilötasolla fenotyyppeihin ja nämä fenotyypit määrää yksilön genotyyppi. Niinpä kelpoisuuden määritelmä yksilön ominaisuudeksi ei ole ongelma vaikka sovellamme kelpoisuutta genotyyppeihin.
Voimme mitata genotyyppistäkelpoisuutta joko absoluuttisesti tai suhteellisesti. Absoluuttinenkelpoisuus on odotettu eloonjäävien jälkeläisten määrä jotka tietyn genotyypin yksilö tuottaa. Suhteellinenkelpoisuus on absoluuttinen kelpoisuus skaalattuna jollakin tavalla. Usein esimerkiksi siten, että suurin kelpoisuus saa arvon 1. Suhteellinen kelpoisuus on usein hyödyllinen evoluution mallintamisessa, mutta absoluuttista kelpoisuutta käyttämällä näemme miten kelpoisuuden käsite on kytköksissä ekologiaan.
Tarkastellaan populaation kasvua yhden sukupolven verran. Olkoon populaatiokoko ajassa t,  . Oletetaan, että tarkasteltavassa lokuksessa on kaksi alleelia ja kuten edellä. Populaatiossa on
. Oletetaan, että tarkasteltavassa lokuksessa on kaksi alleelia ja kuten edellä. Populaatiossa on 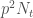 yksilöä joilla on genotyyppi
yksilöä joilla on genotyyppi 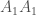 ,
,  genotyypin ja
genotyypin ja  genotyypin
genotyypin  yksilöä. Huomaa, että tiputin termistä alleelia merkitsevän alaindeksin selvyyden vuoksi, lisäksi
yksilöä. Huomaa, että tiputin termistä alleelia merkitsevän alaindeksin selvyyden vuoksi, lisäksi 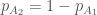 . Käyttäen genotyyppien absoluuttisia kelpoisuuksia, populaatiokoko seuraavassa sukupolvessa on
. Käyttäen genotyyppien absoluuttisia kelpoisuuksia, populaatiokoko seuraavassa sukupolvessa on
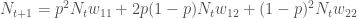 .
.
Tämän voimme kirjoittaa muotoon  , jossa
, jossa  . Termi
. Termi 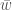 on populaation keskimääräinen kelpoisuus, koska se on genotyyppien kelpoisuuksien summa painotettuna genotyyppien frekvensseillä. Se myös kertoo populaation kasvun yhden sukupolven yli, jos oletamme, että aika on diskreettimuuttuja. Vaikka tämä yhtälö pätee jos sukupolvet eivät ole päällekkäiset (esim. yksivuotiset kasvit), niin usein haluamme ekologiassa kuitenkin mallintaa aikaa jatkuvana muuttujana. Ekologiassa usein populaation kasvua kuvataan termillä
on populaation keskimääräinen kelpoisuus, koska se on genotyyppien kelpoisuuksien summa painotettuna genotyyppien frekvensseillä. Se myös kertoo populaation kasvun yhden sukupolven yli, jos oletamme, että aika on diskreettimuuttuja. Vaikka tämä yhtälö pätee jos sukupolvet eivät ole päällekkäiset (esim. yksivuotiset kasvit), niin usein haluamme ekologiassa kuitenkin mallintaa aikaa jatkuvana muuttujana. Ekologiassa usein populaation kasvua kuvataan termillä
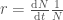 .
.
Jos populaation kasvun on hidasta, niin diskreetti- ja jatkuvamalli ovat likimäärin samat, ja näin löydämme yhteyden populaation kelpoisuuden ja ekologiassa käytetyn populaation kasvun kesken. Siis 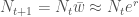 , joten
, joten 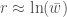 . Ekologiassa käytetty populaation kasvuvauhti on siis populaation keskimääräisen kelpoisuuden luonnollinen logaritmi.
. Ekologiassa käytetty populaation kasvuvauhti on siis populaation keskimääräisen kelpoisuuden luonnollinen logaritmi.
Näin huomaamme, että kelpoisuus siis kytkeytyy ekologiseen teoriaan ja populaation kasvuun. Niinkuin nykyään on tapana sanoa: Fantastista! Kelpoisuudella ei siis ole mitään tekemistä ”vain vahvimmat selviytyvät” tyyppisten sanontojen kanssa. Kyse on siitä miten eri yksilöt vaikuttavat populaation kasvuun. Tietysti yksi osa kelpoisuutta voi olla eloonjääminen, mutta niin ei tarvitse olla.
Kirjallisuutta
Rice, S. H. 2004. Evolutionary Theory: Mathematical and Conceptual Foundations. Sinauer Associates Inc., Sunderland, MA.