Olemme edellisillä kerroilla tarkastelleet populaatiogenetiikan ja ekologian peruskäsitteitä. Nyt pääsemme viimein itse asiaan eli luonnonvalintaan! Luonnonvalinta on tärkein evoluutiomekanismi, vaikka alleelifrekvenssit muuttuvat populaatioissa myös satunnaisajautumisen ja mutaatioiden seurauksena, luonnonvalinta määrää evolutiivisten muutosten suunnan fenotyypin tasolla (lähes kokonaan).
Lähdemme tarkastelemaan luonnonvalintaa populaatiogenetiikan näkökulmasta. Oletamme ensin, että sukupolvet eivät ole päällekkäiset. Lisäksi tarkastelemme ensin tapausta, jossa eri alleeleja kantavien yksilöiden jättämien jälkeläisten määrä seuraavaan sukupolveen riippuu niiden eloonjäämisestä. Kyseessä on siis eloonjäämiseen kohdistuva valinta, luonnonvalinnan ei tietenkään tarvitse kohdistua pelkästään eloonjäämiseen, mutta tämä oletus tekee esimerkistämme yksinkertaisemman.
Tarkastellaan lokusta 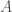
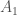

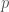
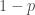

Muistamme edellisestä kirjoituksesta, että populaation keskimääräinen kelpoisuus on 
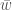
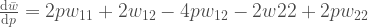
Huomaa, että oletimme tässä, että alleelien kelpoisuudet eivät riipu niiden frekvensseistä.
Kun tarkastelemme alleelifrekvenssien muutoksia, voimme käyttää marginaalisen kelpoisuuden käsitettä. Vaikka valinta kohdistuu genotyyppeihin, oletamme että populaatiossa pariutuminen on satunnaista tarkasteltavan alleelin suhteen. Lasketaan yhteen niiden genotyyppien kelpoisuudet, joissa tarkasteltava alleeli esiintyy, painotettuna näiden genotyyppien frekvensseillä. Olkoon 


Populaation keskimääräinen kelpoisuus marginaalisilla kelpoisuuksilla ilmaistuna on 
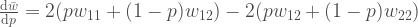
Sijoitetaan marginaaliset kelpoisuudet tähän yhtälöön, niin samme

Marginaalinen kelpoisuus, 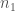

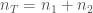




Alleelinfrekvenssin muutos yhdessä sukupolvessa on

Tämä on yleinen muoto alleelifrekvenssien muutokselle. Tarkastellaan omaa esimerkkiämme, jossa meillä oli kaksi eri alleelia. Sijoitetaan
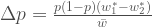
Oletetaan edelleen, että alleelien kelpoisuudet eivät riipu niiden frekvensseistä. Yhdistetään muutos populaation keskimääräisessä kelpoisuudessa ja 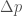

Kun tarkastelemme tätä yhtälöä niin huomaamme muutamia tärkeitä seikkoja: ensinnäkin 

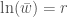
Olemme perimmäisten totuuksien äärellä! Tätä kannattaa pureskella odotellessa seuraavaa blogikirjoitusta.
