Tervehdys jälleen lukijat!
Viimekirjoituksessa oli muutamia kohtia joissa oikaisin muutaman mutkan suoraksi. Tällainen toiminta on kuitenkin paheksuttavaa, joten lienee syytä korjata näitä puutteita.
Muistamme koulusta munkki Mendelin ja hänen kokeensa kasvihybrideillä (Versuche über Pflanzen-Hybriden). Mendelin tulosten merkitystä ei ymmärretty hänen elinaikanaan, mutta ne löydettiin myöhemmin uudelleen.
Kerrataan lyhyesti muistin virkistykseksi mitä Mendelin lait oikein sanovatkaan. Oletamme, että tarkastelemme diplodia eliötä joka lisääntyy suvullisesti. Sanottakoon vielä muutama sana perinnöllisyyden fysikaalisesta perustasta. Tumallisilla otuksilla perimä koostuu usein useasta lineaarisesta DNA-molekyylista. Näitä nimitetään kromosomeiksi. Diploideilla otuksilla on perimässään kaksi kopiota jokaisesta kromosomista, yksi kopio kummaltakin vanhemmalta. Nimitämme näitä vastinkromosomeiksi. Esim. ihmisellä on yhteensä 46 kromosomia, joista 23 olemme perineet isältämme ja 23 äidiltämme. Niinpä kuten aiemmin jo mainittu meillä on kaksi kopiota jokaisesta lokuksesta (jätämme poikkeukset huomiotta).
Ensimmäinen laki – Laki segregaatiosta
Mendelin ensimmäinen laki sanoo että kun yksilö tuottaa sukusoluja jokainen sukusolu saa yhden kopion kustakin lokuksesta (tai alleelista jos niin haluamme)
Toinen laki – Laki riippumattomuudesta
Toinen Mendelin laki sanoo, että alleelit jakautuvat sukusoluihin toisistaan riippumatta. Eli eri lokusten alleelit valikoituvat sukusoluihin itsenäisesti. Esimerkiksi, jos olemme perineet vanhemmalta A 23 kromosomia ja vanhemmalta B 23 kromosomia (ja kuvitellaan, että voimme seurata niitä), omat jälkeläisemme perivät meiltä 23 kromosomia jotka ovat sekoitus A:n ja B:n alkuperäisistä kromosomeista. Toiseen lakiin on olemassa useita tärkeitä poikkeuksia, mutta se jääköön toiseen kertaan.
Mendelin lakien fysikaalinen perusta on kromosomien käyttäytyminen niissä solunjakautumisissa jotka tuottavat sukusoluja. Vastinkromosomit nimittäin asettuvat jakotason eri puolille satunnaisesti. Tätäkin lienee syytä vielä tarkentaa tulevissa kirjoituksissa.
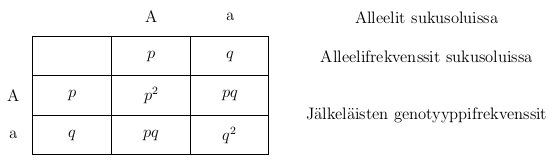
Oheisessa kuvassa on esimerkki risteytyksestä jossa oletan, että tarkasteltavassa lokuksessa on kaksi alleelia, jotka ovat A ja a.

Kun genotyypit A ja a risteytettiin keskenään syntyy genotyyppejä AA, Aa ja aa suhteessa 1 : 2 : 1. Itse koen helpommaksi ajatella asiaa todennäköisyyksien kannalta. Kun Aa genottyyppi tuottaa sukusoluja, Mendelin lakien perusteella sukusolu saa alleelin A todennäköisyydella  ja alleelin a todennäköisyydella . Niinpä tässä tapauksessa jälkeläisen genotyyppi on AA jos se on perinyt alleelin A molemmilta vanhemmiltaan eli
ja alleelin a todennäköisyydella . Niinpä tässä tapauksessa jälkeläisen genotyyppi on AA jos se on perinyt alleelin A molemmilta vanhemmiltaan eli  . Vastaavasti genottyyppi on Aa jos jälkeläinen perii toiselta vanhemmalta alleelin A ja toiselta alleelin a. Tämä voi tapahtua kahdella eri tavalla joten
. Vastaavasti genottyyppi on Aa jos jälkeläinen perii toiselta vanhemmalta alleelin A ja toiselta alleelin a. Tämä voi tapahtua kahdella eri tavalla joten  . Genotyypin aa todennäköisyys lasketaan samoin kuin AA genotyypin.
. Genotyypin aa todennäköisyys lasketaan samoin kuin AA genotyypin.
Tällä kromosomien käyttäytymisellä on useita tärkeitä seurauksia. Se mahdollistaa todennäköisyyslaskennan käytön genetiikassa sekä monia geneettisen analyysin menetelmiä. Sukusolujen taulukointia tai todennäköisyyslaskentaa käyttämällä saamme vanhempien tuottamat jälkeläiset selville millaisessa risteytyksessä tahansa. Mutta mitä tapahtuukaan alleelifrekvensseille populaatiotasolla, kun jokaisessa sukupolvessa lukemattomat yksilöt risteytyvät keskenään ja tuottavat jälkeläisiä?
Oletetaan, että meillä on suuri joukko yksilöitä ja tarkastelleen edellään samaa lokusta, jossa on kaksi alleelia: A ja a. Merkitään A alleelin alleelifrekvenssiä 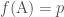 ja a:n vastaavasti
ja a:n vastaavasti  . Alleelifrekvenssit voidaan laskea myös genotyyppifrekvensseistä:
. Alleelifrekvenssit voidaan laskea myös genotyyppifrekvensseistä:  ja
ja  . Alleelifrekvenssien summa on
. Alleelifrekvenssien summa on 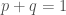 . Nämä ovat siis alleelifrekvenssit nykyisessä sukupolvessa. Nyt kysymme, mitkä ovat alleelifrekvenssit seuraavassa sukupolvessa? Oletan tässä, että tutkimme otuksia, joiden sukupolvet eivät ole päällekkäiset (esim. yksivuotiset kasvit). Tämä tekee esimerkistä yksinkertaisemman, mutta ei ole välttämätön oletus. Oletetaan lisäksi, että yksilöiden tuottamat sukusolut yhdistyvät satunnaisesti.
. Nämä ovat siis alleelifrekvenssit nykyisessä sukupolvessa. Nyt kysymme, mitkä ovat alleelifrekvenssit seuraavassa sukupolvessa? Oletan tässä, että tutkimme otuksia, joiden sukupolvet eivät ole päällekkäiset (esim. yksivuotiset kasvit). Tämä tekee esimerkistä yksinkertaisemman, mutta ei ole välttämätön oletus. Oletetaan lisäksi, että yksilöiden tuottamat sukusolut yhdistyvät satunnaisesti.
Taulukoidaan alleelien mahdolliset tavat yhdistyä kuten edellä. Nyt kuitenkin käytämme alleelifrekvensseja painokertoimina kertomaan kuinka paljon jokaista genotyyppiä on seuraavassa sukupolvessa.
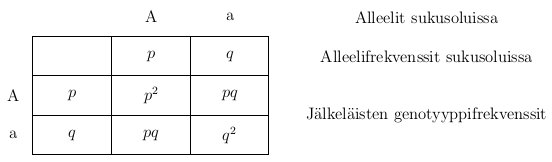
Taulukosta näemme, että seuraavan sukupolven genotyyppifrekvenssit ovat:  ,
,  ja
ja  . Alleelifrekvenssit seuraavassa sukupolvessa ovat siis
. Alleelifrekvenssit seuraavassa sukupolvessa ovat siis  ja vastaavasti
ja vastaavasti  . Käytimme hyväksi edellä kuvattua alleelifrekvenssien laskutapaa genotyyppifrekvensseistä, sekä tietoa
. Käytimme hyväksi edellä kuvattua alleelifrekvenssien laskutapaa genotyyppifrekvensseistä, sekä tietoa 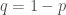 . Huomaamme, että alleelifrekvenssit eivät muuttuneet lainkaan vaan pysyivät samoina! Pelkkä lisääntyminen ei siis muuta alleelifrekvenssejä.
. Huomaamme, että alleelifrekvenssit eivät muuttuneet lainkaan vaan pysyivät samoina! Pelkkä lisääntyminen ei siis muuta alleelifrekvenssejä.
Tätä kutsutaan Hardy-Weinberg laiksi. Jos yksilöt pariutuvat satunnaisesti voimme laskea genotyyppifrekvenssit kun alleelifrekvenssit tunnetaan. Oikeastaan tämä pätee vain jos alleelifrekvenssejä muuttavat voimat eivät vaikuta populaatiossa. Tärkein näistä on luonnonvalinta, muita ovat mutaatiot ja satunnaisajautuminen. Tarkkaan ottaen olemme olettaneet, että tarkasteltava populaatiomme on äärettömän suuri, koska kaikissa rajallisissa populaatioissa alleelien satunnaisotanta seuraavaan sukupolveen muuttaa niiden frekvenssejä aina jonkin verran. Tosin isoissa populaatiossa tämä vaikutus on häviävän pieni. Satunnaisajautuminen onkin merkittävä evolutiivinen voima lähinnä vain erittäin pienissä populaatioissa. Vaikka se onkin tärkeä aihe joillakin populaatiogenetiikan osa-alueilla, tässä blogissa olemme kiinnostuneet evoluutiosta emmekä aio tuhtala aikaamme sen kovin syvälliseen pohtimiseen. Voidaan hyvin kysyä ovatko Hardy-Weinberg yhtälöt kovinkaan tärkeitä, jos monikaan luonnollinen populaatio ei täytä niiden oletuksia? Tässä huolimatta tutkittaessa oikeita populaatioita on todettu, että monet lokukset itseasiassa ovat Hardy-Weinberg tasapainossa. Lisäksi yhtälöt antavat mainion lähtökohdan mutkikkaampien ilmiöiden tarkasteluun.
On myös hyvä huomata pari seikkaa, jotka seuraavat Hardy-Weinberg yhtälöistä. Olen joskus törmännyt kirjoitteluun (netistä löytyy kaikenlaista), jossa on surkuteltu nykyajan surkeaa tilaa ja modernin lääketieteen aiheuttamaa ihmiskunnan geneettistä rappeutumista. Nykyaikaisen lääketieteen ansiostahan moni jos jonkinlaista geneettistä mutaatiota kantava henkilö jää henkiin vaikka vanhoina huonoina aikoja olisi käynyt kalpaten. Vaikka kyseessä on lähinnä angtisten lukiolaisteinien kirjoittelu, johon ei sinänsä tarvitse kiinnittää sen kummempaa huomiota, niin korjaan silti mahdolliset väärinkäsitykset. Edellä kuvatun perusteella voimme todeta, että alleelifrekvenssit pysyvät samoina, jos luonnonvalinta ei niihin vaikuta. Tietysti alleelifrekvenssit voivat muuttua satunnaisotannan seurauksena, mutta isoissa populaatiossa tämä prosessi on niin hidas, että sillä ei juuri ole ihmisen aikaskaalassa merkitystä. Ainoastaan alleelit jotka lisäävät kelpoisuutta voivat luonnonvalinnan vaikutuksesta yleistyä merkittävästi. Niinpä ihmiskunnan geenipoolin tilasta ei tarvitse olla erityisen huolestunut.
On kuitenkin totta, että haitalliset mutaatiot kyllä aiheuttavat populaatioille kelpoisuutta laskevan geneettisen taakan, mutta tämä on kuitenkin eri asia ja lienee oman kirjoituksen aiheensa joskus tulevaisuudessa.
Sillävälin syytä huoleen ei kuitenkaan ole. Otetaan siis rennosti!
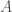
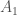

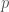
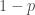


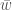
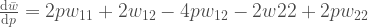




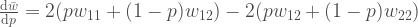

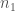

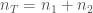





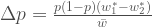
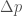



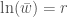

 , on 100 yksilöä. Oletetaan vielä, että nämä yksilöt ovat diploideja (kuten esim. ihmiset ja muut eläimet). Tästä seuraa, että kyseisessä populaatiossa on
, on 100 yksilöä. Oletetaan vielä, että nämä yksilöt ovat diploideja (kuten esim. ihmiset ja muut eläimet). Tästä seuraa, että kyseisessä populaatiossa on 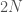 kopiota jokaisesta lokuksesta. Jos populaatiossa on
kopiota jokaisesta lokuksesta. Jos populaatiossa on  alleelin
alleelin 
 muuttuu sukupolvien myötä. Ei oikeastaan mistään sen mystisemmästä. Huomaa, että
muuttuu sukupolvien myötä. Ei oikeastaan mistään sen mystisemmästä. Huomaa, että ![[0, 1]](https://s0.wp.com/latex.php?latex=%5B0%2C+1%5D&bg=ffffff&fg=444444&s=0&c=20201002) . Evoluutio ei kuitenkaan pysähdy jos jokin alleeli yleistyy populaatiossa frekvenssiin 1. Mutaatiot tuottavat uusi alleeleja ja vanhat alleelit korvautuvat uusilla evoluution kuluessa. Luonnonvalinta on tärkein alleelifrekvenssejä muuttava voima. Seuraavaksi selvennämme joitakin käsitteitä, jotka ovat välttämättömiä luonnonvalinnan ymmärtämiseksi.
. Evoluutio ei kuitenkaan pysähdy jos jokin alleeli yleistyy populaatiossa frekvenssiin 1. Mutaatiot tuottavat uusi alleeleja ja vanhat alleelit korvautuvat uusilla evoluution kuluessa. Luonnonvalinta on tärkein alleelifrekvenssejä muuttava voima. Seuraavaksi selvennämme joitakin käsitteitä, jotka ovat välttämättömiä luonnonvalinnan ymmärtämiseksi. . Nyt genotyyppien frekvenssit ovat
. Nyt genotyyppien frekvenssit ovat  ja
ja  . Oikeastaan olisin ennen tätä kirjoitusta voinut selittää tarkemmin miksi näin on. Nyt pyydän lukijoilta malttia ja vain ottamaan tämän annettuna. Tulen selittämään tämän tulevassa kirjoituksessa. Nokkelimmat lukijat ehkäpä muistavat koulusta, että tähän liittyy jotenkin Mendel sekä herrat Hardy ja Weinberg.
. Oikeastaan olisin ennen tätä kirjoitusta voinut selittää tarkemmin miksi näin on. Nyt pyydän lukijoilta malttia ja vain ottamaan tämän annettuna. Tulen selittämään tämän tulevassa kirjoituksessa. Nokkelimmat lukijat ehkäpä muistavat koulusta, että tähän liittyy jotenkin Mendel sekä herrat Hardy ja Weinberg.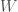 , on yksilön jälkeläisten lukumäärä seuraavassa sukupolvessa. Populaatiogenetiikassa käytämme usein genotyypin kelpoisuutta, joka on kaikkien populaation saman genotyypin yksilöiden keskimääräinen kelpoisuus. Eli genotyypin
, on yksilön jälkeläisten lukumäärä seuraavassa sukupolvessa. Populaatiogenetiikassa käytämme usein genotyypin kelpoisuutta, joka on kaikkien populaation saman genotyypin yksilöiden keskimääräinen kelpoisuus. Eli genotyypin 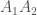 genotyyppinenkelpoisuus,
genotyyppinenkelpoisuus, 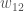 , on kaikkien niiden yksilöiden
, on kaikkien niiden yksilöiden  . Oletetaan, että tarkasteltavassa lokuksessa on kaksi alleelia
. Oletetaan, että tarkasteltavassa lokuksessa on kaksi alleelia 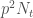 yksilöä joilla on genotyyppi
yksilöä joilla on genotyyppi 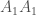 ,
,  genotyypin
genotyypin  genotyypin
genotyypin  yksilöä. Huomaa, että tiputin
yksilöä. Huomaa, että tiputin 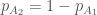 . Käyttäen genotyyppien absoluuttisia kelpoisuuksia, populaatiokoko seuraavassa sukupolvessa on
. Käyttäen genotyyppien absoluuttisia kelpoisuuksia, populaatiokoko seuraavassa sukupolvessa on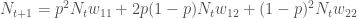 .
. , jossa
, jossa 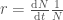 .
.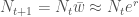 , joten
, joten 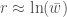 . Ekologiassa käytetty populaation kasvuvauhti on siis populaation keskimääräisen kelpoisuuden luonnollinen logaritmi.
. Ekologiassa käytetty populaation kasvuvauhti on siis populaation keskimääräisen kelpoisuuden luonnollinen logaritmi. ovat sama genotyyppi. Sanokaamme, että alleeli
ovat sama genotyyppi. Sanokaamme, että alleeli 